僼傽乕僕丒僨傿僗僾儗僀偵傛傞峈僀僨傿僆僞僀僾峈懱偺椪彴傊偺墳梡
偼偠傔偵
丂Jerne偺僀僨傿僆僞僀僾乮Id乯丒僱僢僩儚乕僋壖愢埲棃丄惗懱撪偵偍偗傞峈懱嶻惗偼Id-峈Id娫偺憡屳嶌梡偵傛偭偰挷愡偝傟偰偍傝丄帺屓峈懱偼偦偺僶儔儞僗偺攋抅偵傛傝嶻惗偝傟傞壜擻惈偑帵嵈偝傟偰偄傞丅傑偨丄峈Id峈懱偼尦偺峈懱偺僄僺僩乕僾偺撪晹僀儊乕僕傪桳偡傞帠偐傜丄奜棃惈峈尨昡昳拞偺晄弮暔傪娷傑側偄乽埨慡側儚僋僠儞乿條偵墳梡偡傞偙偲偑壜擻偱偁傞丅帠幚丄峈Id峈懱偵傛傞峈尨摿堎揑柶塽墳摎偺桿摫偑帋傒傜傟丄僂僀儖僗峈尨偵懳偡傞峈Id儚僋僠儞偺嶌惉傗丄峈Id峈懱傪梡偄偨B嵶朎惈庮釃傊偺帯椕傕幚嵺偵帋傒傜傟偰偄傞丅偟偐偟丄廬棃偙偆偟偨栚揑偱巊梡偝傟偨儌僲僋儘乕僫儖峈Id峈懱偺懡偔偼儅僂僗丒僴僀僽儕僪乕儅桼棃丄偮傑傝僸僩偵偲偭偰偼堎庬抈敀偱偁傞偨傔丄偙傟傜傪捈愙僸僩偵搳梌偡傞偙偲偼晄弮暔傪娷傑側偄乽埨慡側儚僋僠儞乿傪搳梌偟偨寢壥儅僂僗偺堎庬抈敀傪搳梌偡傞帠偵側傞偲偄偆僷儔僪僢僋僗傪書偊偰偄偨丅帠幚丄峈懱偺搳梌偵傛傝怴偨側傞柶塽斀墳偑庝婲偝傟奺庬傾儗儖僊乕斀墳偑桳奞斀墳偲偟偰婲偙偭偨傝丄帯椕岠壥傕堦帪揑側傕偺偑懡偄帠偑抦傜傟偰偄傞丅傑偨丄屻偵奐敪偝傟偨EB僂僀儖僗丒僩儔儞僗僼僅乕儉偵傛傞儌僲僋儘乕僫儖峈懱偼IgM宆偺峈懱偑懡偔恊榓惈偑掅偄偲偄偆寚揰傪桳偟丄僸僩偺幘姵帯椕偵愊嬌揑偵巊梡偡傞偵偼僷儚乕晄懌偱偁偭偨丅廬偭偰丄僸僩桼棃偺抈敀偱丄偐偮恊榓惈偺崅偄IgG宆偺儌僲僋儘乕僫儖峈Id峈懱偺奐敪偑懸偨傟偰偄偨丅
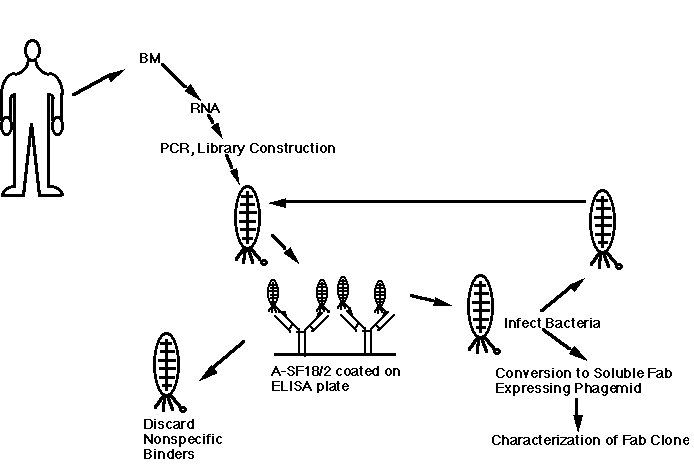
丂杮峞偱偼丄嬤擭奐敪偝傟偨僼傽乕僕丒僨傿僗僾儗僀宆偺僐儞價僫僩儕傾儖峈懱Fab儔僀僽儔儕乕丒僔僗僥儉傪梡偄偰丄僸僩宆偺儌僲僋儘乕僫儖峈Id峈懱傪嶌惉偱偒傞壜擻惈偵偮偄偰弎傋傞丅
1丄僼傽乕僕丒僨傿僗僾儗僀宆偺僐儞價僫僩儕傾儖峈懱Fab儔僀僽儔儕乕丒僔僗僥儉偵傛傞僸僩宆儌僲僋儘乕僫儖峈懱偺敪尰-僋儘乕僯儞僌
丂僐儞價僫僩儕傾儖丒儔僀僽儔儕乕偲偄偆尵梩偼壔妛崌惉偺僕儍儞儖偵偍偄偰傕巊傢傟傞偑丄杮峞偵偍偄偰偼埲屻丄峈懱偺 廳嵔偲寉嵔傪1僙僢僩偯偮偺慻傒崌傢偣偲偟偰帩偮Fab偺儔僀僽儔儕乕傪丄扨偵僐儞價僫僩儕傾儖丒儔僀僽儔儕乕偲屇傇偙偲偵偡傞丅偙偺僔僗僥儉偼堚揱巕岺妛偺庤朄傪婎斦偲偟偰奐敪偝傟丄慜弎偺僴僀僽儕僪乕儅偵偍偗傞僸僩宆峈懱傪嶌惉偱偒側偄偲偄偆寚揰傪崕暈偟丄峏偵戝挵嬠偵偰憹傗偡偙偲偑弌棃傞偨傔僴僀僽儕僪乕儅偲偄偆嵶朎姅傪憹怋偝偣傞偺偵斾傋帪娫偲僐僗僩傪尭傜偡帠傪壜擻偲偟偨丅條乆側曽朄偑妋棫偝傟偮偮偁傞偑丄尰嵼偺偲偙傠僗僋儕僢僾僗尋媶強乮La Jolla, CA. USA乯偺曽朄偲丄働儞僽儕僢僕峈懱尋媶強乮Cambridge.UK乯偺曽朄偑2戝庡棳偱偁傞丅僗僋儕僢僾僗偺僔僗僥儉偼丄僸僩偺崪悜側偳傛傝mRNA傪拪弌偟cDNA傪崌惉屻丄廳嵔偍傛傃寉嵔堚揱巕傪奺乆悢慻偺僾儔僀儅乕丒僙僢僩傪梡偄偰RT-PCR偱憹暆偟丄廳嵔偼Xho1, Spe1丄寉嵔偼Xba1, Sac1偲偄偆惂尷峺慺偱愗抐屻偵弴師pComb3傑偨偼pComb3H偲偄偆儀僋僞乕偵慻傒崬傒儔僀僽儔儕乕傪峔抸偡傞丅堦曽偺働儞僽儕僢僕偺僔僗僥儉偱偼丄廳嵔偲寉嵔傪RT-PCR偱憹暆偡傞偲偙傠傑偱偼摨條偩偑丄偦偺屻廳嵔偲寉嵔傪儕儞僇乕偵偰愙懕屻偵2nd PCR傪峴側偄丄Sfi1, Not1偲偄偆惂尷峺慺偵偰愗抐屻偵堦妵偟偰pCANTAB5E偲偄偆儀僋僞乕偵慻傒崬傒儔僀僽儔儕乕傪峔抸偡傞丅偙傟傜偺儔僀僽儔儕乕偵M13宯偺僿儖僷乕丒僼傽乕僕傪姶愼偝偣傞帠偵傛傝僼傽乕僕昞柺偵堦慻偺Fab傪昞弌偡傞僼傽乕僕丒僨傿僗僾儗僀宆偺僐儞價僫僩儕傾儖丒儔僀僽儔儕乕偲側傞丅偦偺屻偺峈尨摿堎揑側Fab僋儘乕儞傪儔僀僽儔儕乕傛傝慖戰-擹弅偡傞夁掱偼Panning偲屇偽傟偰偄傞丅捠忢偼ELISA僾儗乕僩摍偵屌憡壔偟偨峈尨偵僼傽乕僕丒僨傿僗僾儗僀宆偺僐儞價僫僩儕傾儖丒儔僀僽儔儕乕傪堦掕帪娫斀墳偝偣偨屻丄旕摿堎揑寢崌傪愻偄彍偒丄摿堎揑偵寢崌偟偨僼傽乕僕偺傒傪巁偱梟弌偟丄嵞搙戝挵嬠偵姶愼偝偣憹暆偟偦偙偐傜僼傽乕僕傪拪弌偟丄師偺儔僂儞僪偺峈尨偵斀墳偝偣傞丅偙偺夁掱傪捠忢偼3乣6儔僂儞僪孞傝曉偡帠偵傛傝丄峈尨偵摿堎揑偵寢崌偡傞Fab傪帩偮僼傽乕僕傪擹弅偱偒傞丅堦斒偵1儔僂儞僪偺Panning偱峈尨偵摿堎揑偵寢崌偡傞Fab傪帩偮僼傽乕僕傪偍傛偦100攞偵擹弅偱偒傞偲偝傟偰偄傞丅Panning偺嵟廔儔僂儞僪屻丄僗僋儕乕僯儞僌偺偨傔偺壜梟惈Fab傪嶌惉偡傞偑丄偙偙偱傕傑偨2偮偺僔僗僥儉偼堎側偭偰偄傞丅僗僋儕僢僾僗偺僔僗僥儉偱偼丄GeneIII偲偄偆堚揱巕晹暘乮廳嵔堚揱巕傪戝挵嬠偺嵶朎暻撪枌偵屌掕偡傞Coat protein III偲偄偆抈敀傪僐乕僪偡傞乯傪Nhe1, Spe1偲偄偆惂尷峺慺偵偰愗抐屻偵僙儖僼丒儔僀僎乕僔儑儞偝偣傞帠偵傛傝丄壜梟惈偺Fab傪敪尰偡傞僾儔僗儈僪偵揮姺偱偒傞丅堦曽偺働儞僽儕僢僕偺僔僗僥儉偱偼戝挵嬠傪僲儞僒僾儗僢僒乕姅偵慻傒懼偊傞帠偵傛傝壜梟惈偺Fab傪敪尰偱偒傞丅偙偺傛偆偵偟偰嶌惉偟偨壜梟惈Fab梟塼傪梡偄偰捠忢偼ELISA摍偵偰峈尨摿堎揑側Fab傪僗僋儕乕僯儞僌偡傞丅椙偄Fab僋儘乕儞偑摼傜傟傟偽戝検攟梴屻偵惛惢偟偦偺恊榓惈丄摿堎惈丄岎嵆斀墳惈側偳偑挷傋傜傟傞丅偙偺椉幰偺僔僗僥儉偺桪楎偵娭偟偰偼挊幰偼慜幰偺僔僗僥儉偟偐巊梡宱尡偑側偄偨傔尵媦偱偒側偄丅彯丄屻幰偼Pharmacia Biotech傛傝僉僢僩偲偟偰巗斕偝傟偰偄傞偑丄慜幰偵娭偟偰偼崱偺偲偙傠庤偵擖傟傞偵偼僗僋儕僢僾僗尋媶強偵捈愙僐儞僞僋僩偟峸擖偡傞偟偐側偄丅
2丄僸僩宆儌僲僋儘乕僫儖峈懱偺崱傑偱偺揔梡
丂僸僩宆峈懱傪幚嵺偵僸僩偵搳梌偱偒傞偲偄偆棙揰偐傜丄姶愼徢偺帯椕摿偵尰嵼偁傞栻嵻偑尷傜傟偨岠壥偟偐側偄僂僀儖僗幘姵偵懳偟偰拞榓峈懱傪搳梌偡傞丄偮傑傝偼庴摦柶塽偺曽岦偐傜惙傫偵尋媶偑姪傔傜傟偰偄傞丅崱傑偱偺偲偙傠丄HIV丄扨弮僿儖儁僗僂僀儖僗丄悈摋懷忬醰怾僂僀儖僗丄RS僂僀儖僗丄EB僂僀儖僗丄僒僀僩儊僈儘僂僀儖僗丄杻怾僂僀儖僗丄晽怾僂僀儖僗丄僨儞僌僂僀儖僗丄僄儃儔僂僀儖僗丄儘僞僂僀儖僗摍偵懳偡傞僸僩宆峈懱偑僋儘乕僯儞僌偝傟丄偦偺堦晹偼婛偵椪彴墳梡偑巒傑偭偰偄傞丅偙傟傜僂僀儖僗偵懳偡傞峈懱偼偦偺恊榓惈偩偗偱偼側偔丄偦偺拞榓妶惈偺嫮偝偑廳梫偲側傞丅
丂堦曽丄帺屓柶塽幘姵偵偍偗傞帺屓峈懱偵娭偟偰傕條乆側幘姵偺姵幰桼棃偺僸僩宆儌僲僋儘乕僫儖帺屓峈懱偑僋儘乕僯儞僌偝傟偰偄傞丅峈DNA峈懱丄峈妀峈懱丄峛忬態儁儖僆僉僔僟乕僛傗僒僀儘僌儘僽儕儞偵懳偡傞峈懱丄儈僩僐儞僪儕傾帺屓峈尨PDC-E2偵懳偡傞峈懱丄pANCA乮岲拞媴嵶朎幙峈懱乯丄U1
RNA娭楢A抈敀偵懳偡傞峈懱丄峈傾僙僠儖僐儕儞丒儗僙僾僞乕峈懱丄峈僇儖僕僆儕僺儞峈懱丄峈SS-A峈懱摍偑曬崘偝傟偰偄傞丅偟偐偟側偑傜丄帺屓峈懱傪捈愙僸僩偵搳梌偟偰傕懄帯椕偲側傞栿偱偼側偄偺偱丄椪彴墳梡偲側傞偲傕偆堦傂偹傝擇傂偹傝偑昁梫偲側偭偰偔傞丅帺屓峈懱偺傾僢僙僀宯偺梲惈僐儞僩儘乕儖偵偡傞摍丄恌抐傊偺墳梡偼婔偮偐壜擻偱偁傠偆丅傑偨丄偙偺庬偺尋媶偑惙傫偵峴側傢傟傟偽丄嵟廔揑偵偼僸僩僎僲儉偺慡夝愅偲偄偆敪憐偵帡偨丄慡偰偺帺屓峈尨偍傛傃帺屓峈懱偺僋儘乕僯儞僌偲偄偆栚昗傕払惉偱偒傞偐傕偟傟側偄丅
3丄僔僃乕僌儗儞徢岓孮偵偍偗傞Vg堚揱巕嵞峔惉
丂偙偙偐傜丄摉壢偵偍偗傞僔僃乕僌儗儞徢岓孮乮SS乯偵偍偗傞儕僂儅僩僀僪場巕乮RF乯偵懳偡傞嫟捠Id偵娭偡傞抦尒偵娭偟偰弎傋傞丅傑偢丄SS偵偼埆惈儕儞僷庮傗M抈敀寣徢摍偺儕儞僷憹怋惈昦曄偺崌暪偑懡偄帠偑抦傜傟偰偄傞丅崌暪偡傞埆惈儕儞僷庮偼斾妑揑掅埆惈搙乮Indolent乯偵懏偡傞Marginal
zone B嵶朎儕儞僷庬偑懡偄偑丄偦偺懠偺慻怐宆偺傕偺傕曬崘偝傟偰偍傝丄偦偺婋尟搙偼曬崘偵傛傝傑偪傑偪偩偑悢廫攞偲峫偊傜傟傞丅
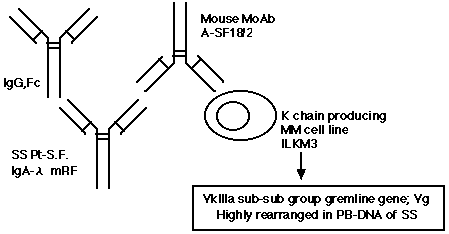
丂変乆偼SS姵幰桼棃偺4庬椶偺儌僲僋儘乕僫儖RF偵懳偟偰廬棃偺儅僂僗丒僴僀僽儕僪乕儅朄偵傛傝儌僲僋儘乕僫儖峈Id峈懱傪嶌惉偟偨丅偦偺寢壥丄SS姵幰偱偼崅棪偵嫟捠Id傪桳偟偰偄傞帠偑暘偐偭偨丅側偐偱傕丄IgA-兩宆M抈敀寣徢丄崅兞僌儘僽儕儞寣徢惈巼斄丄夁擲鈌徢岓孮傪桳偡傞姵幰SF桼棃偺A-SF18/2偼丄a乯寬忢幰偵斾傋丄SS姵幰偺寣惔偍傛傃枛徑寣儕儞僷媴偵崅棪偵斀墳偟偨丅b乯內嵔嶻惗懡敪惈崪悜庮嵶朎姅ILKM3偵斀墳偟偨丅偦偙偱丄ILKM3偺柶塽僌儘僽儕儞寉嵔V內堚揱巕偺墫婎攝楍傪挷傋偨偲偙傠丄V內IIIa
sub-subgroup偵懏偡傞Vg堚揱巕偲95.8%偺嫟捠惈偑擣傔傜傟偨丅
丂廬偭偰丄SS姵幰偵偍偄偰Vg堚揱巕傑偨偼椶帡偺V內III subgroup堚揱巕偑崅棪偵曐懚偝傟側偑傜巊梡偝傟偰偄傞壜擻惈偵偮偄偰専摙偟偨丅偦偺寢壥丄SS姵幰枛徑寣儕儞僷媴偵偍偄偰丄V內III
subgroup堚揱巕偑崅棪偵嵞峔惉偟擣傔傜傟偨丅傑偨丄Vg堚揱巕偺嵞峔惉僋儘乕儞偺妱崌偵娭偟偰丄Bridges傜偺枬惈娭愡儕僂儅僠偵偍偗傞曬崘乮16%乯偵偵斾傋丄SS偱偼崅棪乮40%乯偲崅棪偱偁偭偨丅峏偵丄嵞峔惉偟偰偄偨Vg僋儘乕儞偺Germline偲偺憡摨惈偼丄摨堦姵幰偵偍偄偰傕98.6%,
95.8%偲堎側偭偰偍傝丄庡偵曄堎偑CDR3椞堟偵懚嵼偟偰偄偨乮Ongoing mutation乯丅埲忋傛傝丄SS姵幰偺枛徑寣儗儀儖偱Vg堚揱巕偑丄崅棪偵僒僓儞僽儘僢僩偱専弌偝傟傞掱搙偺曄堎偱曐懚偝傟側偑傜傕嵞峔惉偟偰偍傝丄偐偮Ongoing偱偁傞帠偑丄嫟捠Id偑崅棪偵擣傔傜傟丄偦偙偐傜儕儞僷憹怋惈昦曄偑敪徢偟傗偡偄堦偮偺梫場偲峫偊傜傟偨乮恾1乯丅
丂偟偐偟側偑傜丄A-SF18/2偺斀墳偡傞僄僺僩乕僾偑丄廳嵔偲寉嵔偺偄偢傟偐丄傕偟偔偼偦偺椉幰偱偁傞偺偐丄偦偺偳偺晹暘偱偁傞偺偐摍偺媈栤偑巆偭偰偄偨丅
4丄僔僃乕僌儗儞徢岓孮偵偍偗傞僀僨傿僆僞僀僾丒僱僢僩儚乕僋
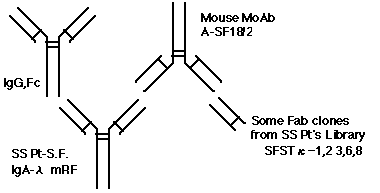
丂愭偺媈栤傪夝寛偡傞偨傔丄僐儞價僫僩儕傾儖丒儔僀僽儔儕乕傪梡偄偰丄A-SF18/2偵斀墳偡傞Fab僋儘乕儞傪摼傞帠傪栚揑偵埲壓偺幚尡傪峴側偭偨丅
丂傑偢丄枬惈娭愡儕僂儅僠偲擇師惈SS偺姵幰乮屻偵埆惈儕儞僷庮傪敪徢偟巰朣丄峈妀峈懱亊160 Speckled丄RF亊10000丄ELISA偵偰寣惔偼A-SF18/2偵嫮梲惈偵斀墳乯偺崪悜傛傝RNA傪拪弌偟丄慜弎偺庤朄偱僐儞價僫僩儕傾儖丒儔僀僽儔儕乕傪嶌惉偟偨丅屌憡壔偟偨A-SF18/2偵懳偟偰寁6儔僂儞僪偺Panning傪峴側偄丄A-SF18/2偵懳偟偰斀墳偡傞Fab僋儘乕儞傪ELISA偵偰僗僋儕乕僯儞僌偟偨乮恾2乯丅
丂偦偺寢壥丄寁5僋儘乕儞偺Fab丟SFST內-1,2,3,6,8偑摼傜傟偨乮恾3乯丅偙偺偆偪SFST內-3偼A-SF18/2偲嫮偄恊榓惈傪桳偟偰偄偨偑丄僆僶儖僽儈儞,
僸僩IgG-Fc, 儅僂僗IgG-Fc偲傕岎嵆斀墳惈傪帵偟偨丅堦曽丄SFST內-1,2,6,8偼A-SF18/2偲偺恊榓惈偼庛偐偭偨偑岎嵆斀墳惈偼柍偐偭偨丅傑偨5偮偺Fab僋儘乕儞偺慡偰偑廳嵔堚揱巕傪桳偟偰偄偨偑丄寉嵔堚揱巕偼SFST內-3偲6偺傒偵偁傝丄1,2,8偼寉嵔偑側偐偭偨丅廳嵔偺傒偱傕恊榓惈傪擣傔偨僋儘乕儞偑偁傞偙偲偐傜丄A-SF18/2偼廳嵔偍傛傃寉嵔偺偳偪傜偐堦曽偵偱傕寢崌偟偆傞帠偑帵嵈偝傟偨丅嵟傕嬤偄Germline堚揱巕偼丄廳嵔偼SFST內-1偑DP-77/WHG16丄2偑DP-47/V3-23丄3偑DP-38/9-1丄6偑DP-47/V3-23丄8偑DP-50/hv3019b9偱偁傝丄2偲6偑嫟捠偱丄傑偨慡偰VH3
subgroup偱偁偭偨丅堦曽丄寉嵔偺Germline偼SFST內-3偍傛傃6偑嫟偵嵟傕嬤偄偺偑DPK22/A27偱丄2斣栚偵嬤偄傕偺偑慜弎偺Vg偱偁偭偨丅傑偨丄峈懱偺峈尨摿堎惈偼CDR3椞堟偑廳梫偲偝傟偰偄傞偨傔丄奺僋儘乕儞偺CDR3椞堟傪斾妑偟偨偲偙傠丄廳嵔偱偼SFST-1丟DRGMYSSGT丄2丟QGSGWHHDA丄3丟ARLGYGRAL丄6丟DPTYYGDFGG丄8丟DDGGS偱偁傝丄寉嵔偱偼3丟QNYGNSP丄6丟QQYSRSP偱柧傜偐側嫟捠惈偼擣傔傜傟側偐偭偨丅廬偭偰丄A-SF18/2偼柶塽僌儘僽儕儞偺摿掕偺傾儈僲巁攝楍偱偼側偔丄壗傜偐偺棫懱峔憿傪擣幆偟偰偄傞壜擻惈偑帵嵈偝傟偨丅SFST內-3偺傛偆偵嫮偄恊榓惈傪桳偡傞僋儘乕儞偑傕偭偲懡偔摼傜傟傟偽丄寢崌偡傞僄僺僩乕僾傪峏偵偼偭偒傝偲摨掕偱偒傞偲峫偊傜傟丄尰嵼摉尋媶傪宲懕拞偱偁傞丅
丂彮側偔偲傕丄僼傽乕僕丒僨傿僗僾儗僀宆偺僐儞價僫僩儕傾儖丒儔僀僽儔儕乕偐傜丄峈Id峈懱偵寢崌偡傞峈懱Fab僋儘乕儞丟峈-峈Id峈懱偑摼傜傟傞帠偑柧傜偐偲側偭偨丅
5丄僸僩宆儌僲僋儘乕僫儖峈僀僨傿僆僞僀僾峈懱偺崱屻偺墳梡
丂傑偢丄崱傑偱偺偲偙傠尷傜傟偨椺偱偟偐帵偝傟偰偄側偄Id僱僢僩儚乕僋傪徹柧偡傞昁梫偑偁傞偲巚傢傟傞丅椺偊偽崱夞偺変乆偺Fab僋儘乕儞偼丄RF偵懳偡傞峈Id峈懱偵懳偡傞峈懱偱偁傞偐傜丄RF偲摨條偺峈懱妶惈傪桳偟偰偄傞壜擻惈偑偁傞丅帠幚SFST內-3偼僸僩IgG-Fc偲傕岎嵆斀墳惈傪桳偟偰偄偨丅偟偐偟丄懠偺僋儘乕儞偼RF妶惈傪帵偝側偐偭偨帠偐傜丄Id偺僱僢僩儚乕僋偼偦傟傎偳尩枾側傕偺偱偼側偔丄條乆側峈懱偑岎嵆斀墳惈偲偄偆晹暘偱儖乕僘偵宷偑偭偰偄傞壜擻惈偑偁傞丅
丂傑偨丄帺屓柶塽幘姵姵幰偺峈懱嶻惗僋儘乕儞偼捠忢億儕僋儘乕僫儖偵妶惈壔偝傟偰偍傝丄偦偺拞偵偼條乆側峈尨偵岎嵆斀墳偡傞僋儘乕儞傕摉慠懡偔娷傑傟偰偍傝丄廬偭偰崱夞偺傛偆側庤朄偱僸僩宆儌僲僋儘乕僫儖峈Id峈懱傪摼傗偡偄壜擻惈偑偁傞丅堦曽丄帺屓柶塽幘姵傪桳偟側偄姶愼徢姵幰偵偍偗傞堎庬抈敀偵懳偡傞峈懱偵娭偟偰偼偳偆偩傠偆偐丅椺偊偽丄HIV偺gp120偵懳偡傞僸僩宆峈懱偵懳偟偰峈Id峈懱偑摼傜傟傟偽丄gp120偺撪晹僀儊乕僕傪帩偮儚僋僠儞偲偟偰巊梡偱偒傞栿偩偑丄崱傑偱偺偲偙傠偦偺傛偆側曬崘偼尒摉偨傜側偄丅
丂廬偭偰丄僸僩宆儌僲僋儘乕僫儖峈Id峈懱傪梡偄傞帠偵傛傝丄崱傑偱偺姶愼徢偵懳偡傞庴摦柶塽庡懱偺峈懱棙梡偵戙傢傝丄儚僋僠儞偲偟偰梡偄傞擻摦柶塽偲偟偰偺棙梡傊揮姺偱偒傞壜擻惈偼栜榑偁傞丅偟偐偟丄偦偺偨傔偵偼峈懱偑摼傜傟側偗傟偽側傜側偄偑丄偦偙偵偼傑偩晄柧側晹暘傕偁傝崱屻偺専摙壽戣偱偁傞丅
偍傢傝偵
丂僐儞價僫僩儕傾儖丒儔僀僽儔儕乕傪梡偄偨敪尰-僋儘乕僯儞僌偵傛傝峈Id峈懱偺僄僺僩乕僾傪摨掕偱偒丄Id僱僢僩儚乕僋傗帺屓峈懱嶻惗偺婡彉偵懳偡傞棟夝傪怺傔傜傟傞偱偁傠偆丅峈Id峈懱傪梡偄偨儚僋僠儞奐敪偺壜擻惈偵娭偟偰偼丄峏側傞搘椡偑昁梫偲巚傢傟傞丅
{kind=link}
{kind=link}
{kind=link}